


¿Qué es eso de las especies? Parte 1: los organismos “fáciles”
A ver: va con mallas azules, vuela con una capa roja y se peina con raya. Está claro: es Supermán. Acabamos de hacer una clasificación. Clasificar un individuo consiste en asignarle a una clase, que debe estar definida por un conjunto de características diagnósticas. Dentro de la clase todos los individuos serán esencialmente iguales, es decir, cumplirán esas características.
Por ejemplo, vemos un piolín por el campo y observamos que mide un par de palmos, que tiene pinta de urraca con la mitad de la cabeza negra y la cola y alas azules. Pues con eso se acabó el problema: es un rabilargo (Cyanopica cyanus). Asumimos que la clase “rabilargo” define individuos que, sin excepciones, poseen las características mencionadas y que ese conjunto de características no se repite en otras clases.
En el problema que nos ocupa, las clases serán las especies (en biología hay una categoría taxonómica que se llama Clase, pero no me refiero a eso). Como he comentado, cada especie debe definirse mediante unas características que permitirán separarla de las demás. Pero antes de eso debemos definir qué es una especie, o dicho de otra forma, que significado tiene la clase “especie”.
La definición clásica (BSD, Biological Species Definition) es la propuesta por Ernst Mayr en el libro Systematics and the Origin of Species publicado en 1942:
“groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups”.Interpretado libérrimamente, un individuo pertenece a una especie si puede reproducirse con los demás individuos de esa especie generando a su vez descendientes que pueden seguir el proceso por ser también fértiles. Y podremos decir que una especie se diferencia de otra especie si los organismos de cada una no pueden fecundarse o, al menos, si los descendientes son estériles.
La definición se apoya en la diferenciación genética: dos individuos que comparten esencialmente el mismo genotipo pertenecen a la misma especie; la similitud genotípica se refleja en la capacidad de fecundación y de tener descendencia. La distancia genotípica hace que la fecundación sea imposible por causas diversas: etológicas o propiamente genéticas (por ejemplo, por tener diferente número de cromosomas).
El ejemplo que comentábamos en las entradas anteriores del perro y del lobo estaría claro: son una misma especie a pesar de que existen diferencias externas ya que la distancia genética entre ambos no es aún suficiente como para evitar la fertilización y los descendientes fértiles. En cambio, los humanos y los chimpancés no somos la misma especie a pesar de que nuestros genotipos son bastante similares: nosotros tenemos 46 cromosomas y nuestros primos 48.
Tenemos por tanto una herramienta para separar especies que, además, tiene sentido evolutivo. Abajo vemos un gráfico que reflejaría el proceso estándar.
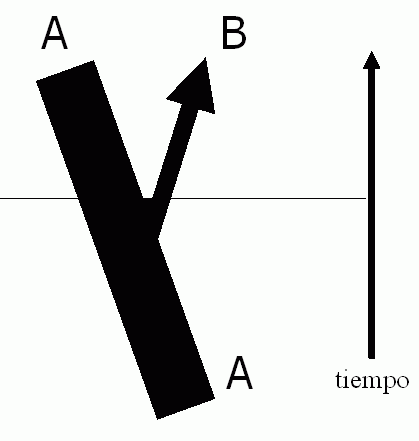
Comenzamos con una especie, A, formada por un conjunto de poblaciones (grupos de individuos conectados genéticamente de forma real, no sólo potencial, por vivir en la misma área). En cierto momento, una o más de esas poblaciones se separa genéticamente del grupo principal (normalmente por separación física). La ausencia de intercambio genético hace que esos dos grupos evolucionen independientemente a partir de entonces. La línea horizontal representa el punto de no retorno, cuando esa divergencia evolutiva ya es tan grande que las dos ramas no son interfértiles: ha surgido la especie B.
De este esquema deducimos un par de cosas interesantes.
La primera es que las especies son entidades no permanentes. El catálogo de especies en la Tierra varía según donde pongamos la línea horizontal de la figura. Esa línea representa un corte en un continuo árbol filogenético cuyas ramas varían con el tiempo, algunas surgen, otras desaparecen, incluso la especie A deberá llamarse de forma distinta dentro de un tiempo porque habrá variado sustancialmente respecto a sus ancestros.
La segunda es que la prueba de interfertilidad no puede hacerse en la práctica. La definición de Mayr es más una construcción lógica razonable que una herramienta de diagnóstico. Y como veremos en otra entrada, hay muchos organismos que se resisten a someterse a ella porque la BSD sólo es aplicable a organismos con reproducción sexual y en este mundo hay cosas mucho más raras.
Para facilitarnos la vida, la solución ha sido asumir que las diferencias genotípicas van a tener un reflejo en los caracteres observables del bicho o planta con lo que bastaría con dar unos buenos criterios morfológicos para resolver el problema (eso hemos hecho con Supermán y con el rabilargo). En la realidad, casi siempre se trabaja así con animales y vegetales “superiores”: la morfoespecie se asimila a la especie biológica. En paleontología no cabe otra opción.
El uso de caracteres morfológicos funciona razonablemente bien en muchos casos como, por ejemplo, en vertebrados. Aún así, hay casos que pueden llevar a confusión, como especies polimorfas, diformismo sexual… Pero funciona sólo regular en muchos grupos y no funciona en absoluto en otros que comentaremos en otro momento.
Por tanto, el concepto de especie es ante todo utilitario y adaptado a la escala temporal humana, donde la evolución apenas es perceptible. La definición básica es razonable pero no deberíamos tomarnos al pie de la letra que sea aplicable a todos los organismos porque, de hecho, no lo es. Su mayor problema es que la aplicación de la BSD de Mayr necesita criterios que son mecanismos potenciales o aluden a relaciones no comprobables por lo que, aunque aclara el concepto, lleva el problema práctico a un punto muerto del que sólo puede salirse con soluciones de compromiso donde se sacrifica la coherencia en aras de la utilidad.
Nota: este tema entra plenamente en mi campo de incompetencia, agravado por la necesaria brevedad del post. Hay libros enteros sobre el asunto de la especiación, resumir lo esencial de la cuestión en dos folios es un atrevimiento que espero disculpen y corrijan en los comentarios.


{kind=link}
1 comentarios:
Angel, has puesto a tiro un chiste fácil. Habría que preguntar a Lois Laine si el Homo sapiens kriptonensis está emparentado con el rabilargo.
Perdón, pero tenía que decirlo.
Entiendo que dos especímenes pertenecen a la misma especie si pueden tener descendencia fértil entre ellos. Que, siendo del mismo género, es difícil saberlo (dejando análisis genéticos aparte) porque habitualmente no comparten el mismo hábitat, y sólo una nueva migración, o la intervención humana, puede volver a juntarlos.
Pero, ¿qué pasa con las especies asexuadas? Tal vez sea esta una pregunta absurda, porque si este tema entra plenamente en tu campo de incompetencia, no te cuento cuál es la mía: 0 ó -1.
Publicar un comentario