Se conocen aproximadamente 1.4 millones de especies de organismos vivos (obviaremos aquí el debate sobre qué significa “especie”). Se estima que esta cifra representa el 15% del número real y algunas otras estimaciones lo sitúan en apenas un 2%. Peter H. Raven y Edward O. Wilson comentaron hace ya 15 años algunos datos al respecto [DOI: 10.1126/science.258.5085.1099]; los comentarios entre paréntesis son míos:
- En la década de los 80 fueron descubiertas tres nuevas familias de plantas con flores en Centroamérica.
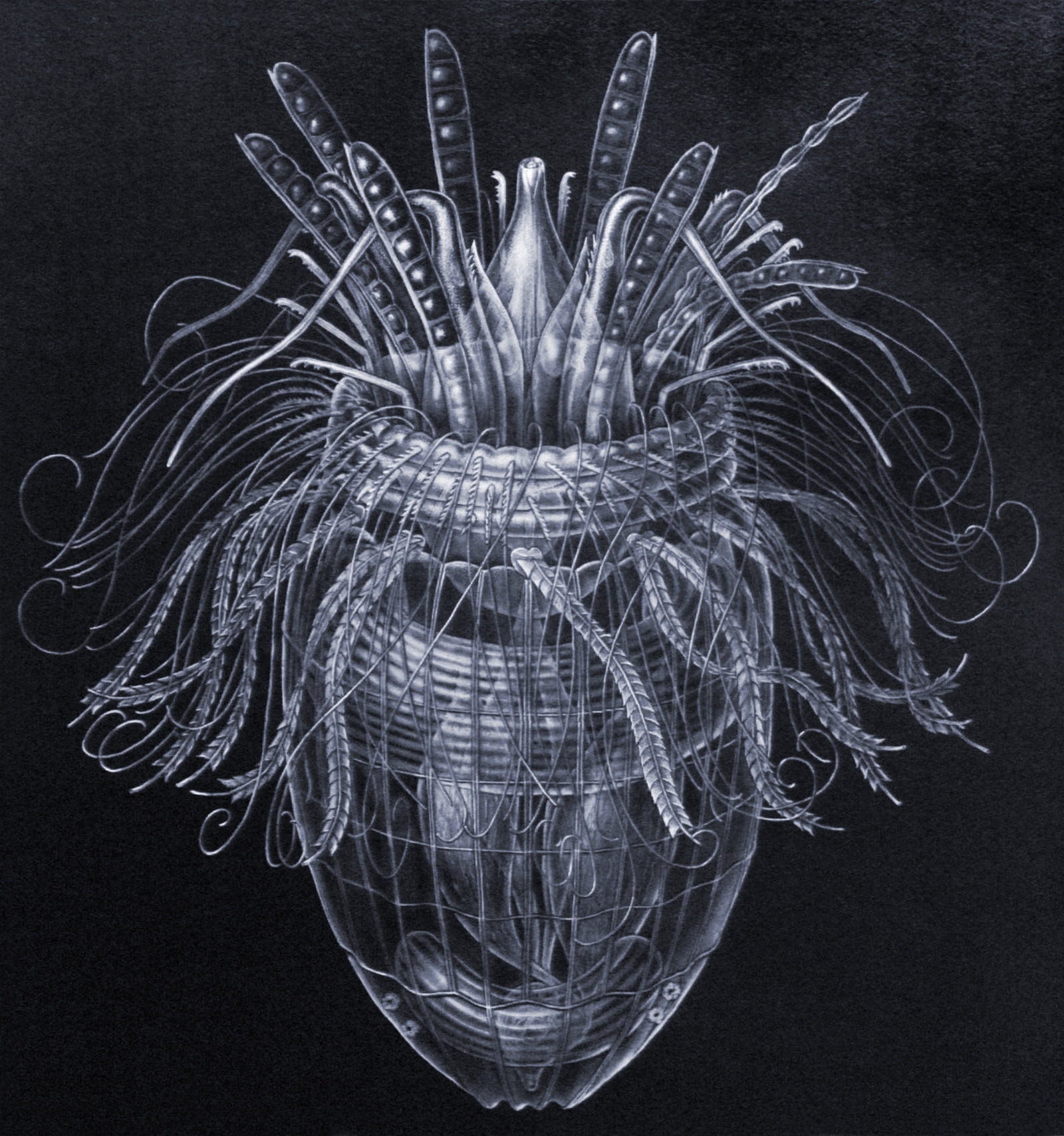
- El filum animal más reciente (Loricifera) fue descrito en 1983 (recordemos que filum representa un nivel taxonómico muy alto, como los Cordados, Artrópodos o Moluscos y que se basa en diseños anatómicos exclusivos).
- La gran mayoría de los insectos del dosel arbóreo tropical, probablemente más del 90% en algunos grupos, aún es desconocido.
- Se han descrito unas 69000 especies de hongos pero se estima que hay un mínimo de 1.5 millones.
- Los microbiólogos reconocen unas 4000 especies de bacterias pero se supone que son sólo una ínfima parte de la realidad. Un gramo de suelo en un bosque puede contener 4000-5000 especies pero cuyas condiciones de cultivo se desconocen; lo mismo ocurre en un gramo de sedimento marino, sólo que las especies son diferentes.

Pliciloricus enigmatus, una de las especies del nuevo filum Loricifera
Estos datos están desfasados por la fecha de publicación. Por ejemplo, desde entonces se han descubierto dos nuevos filum: Cycliophora y Micrognathozoa. La situación general, sin embargo, no ha cambiado: el progreso hacia el conocimiento de la prodigiosa biodiversidad de la Tierra ha sido y es muy lento. En ese artículo Raven y Wilson urgían a reforzar el inventario de especies, el 80% de las cuales está en los trópicos, las zonas más afectadas hoy por la destrucción de habitats.
Podríamos suponer que no es mal momento para ello. “Biodiversidad” es la palabra de moda (en dura competición con eso del “desarrollo sostenible”) y cabría esperar recursos e iniciativas para su estudio. Pero no es exactamente así. La biodiversidad se ha convertido en un concepto propagandístico que sólo se financia cuando es titular de la prensa. Pero bueno, ya me voy por las ramas, volvamos al tema.
Las especies se descubren cuando un especialista las encuentra, analiza, compara con lo ya conocido… y las describe siguiendo un protocolo bien establecido. Deben guardarse varios ejemplares incluyendo aquel en el que se ha basado la descripción y que se denomina holotipo. Esos ejemplares residen físicamente en colecciones de esas que parecen decimonónicas: cajones y archivadores llenos de carcasas de insectos, pliegos de herbario con plantas secas entre sus hojas… Estas colecciones atesoran millones de especímenes (por ejemplo, un millón en el RJB, 6 millones en el MOBOT) que pueden ser pedidos, prestados y estudiados por especialistas de todo el mundo. En algún lugar hay pliegos con especies ya extintas, o variedades de plantas antes cultivadas y ahora ya olvidadas. Un tesoro de diversidad genética y de información de primera mano.

Holotipo de Carduus modestii Tamamsch recolectado en Ulan-Bator, Mongolia, en 1927, actualmente depositado en el Missouri Botanical Garden (pinche aquí para verlo en alta resolución, 1.8 Mb).

Etiquetas del ejemplar anterior, obsérvese la llamada de atención: Typus!
Pero esa es la parte menos popular de la taxonomia y, curiosamente, los fondos no van a ampliar las colecciones. Scott Miller comenta:
“It is ironic that, just as the U.S. National Science Foundation increases funding for biodiversity research, many states are threatening to discontinue support for their collections”
En Challenges and opportunities in understanding and utilisation of African insect diversity (Miller, S.E, and and Rogo, L.M., 2001, Cimbebasia, 17:197-218) da alguna razón práctica para ello:
“Yet, there is urgent need for basic information on insect diversity for pest management related to plant, livestock and human health, as well as conservation and environmental management.”
¿Qué pasa? ¿Dónde está realmente el estudio de la biodiversidad?
Peter Raven y Edward Wilson vuelven a la carga años después y, con Quentin D. Wheeler, dicen que los fondos que nominalmente están destinados a la taxonomía se dedican de forma mayoritaria a análisis de ADN y a la construcción de filogenias moleculares. Mientras tanto, insisten, cientos de especies están amenazadas de extinción. Y su conocimiento se detiene, añado yo.
En mi opinión de no-especialista en nada, es un problema de sexy-science: hoy molan las filogenias moleculares pero no el trabajo de campo. Las primeras forman parte de la “gran ciencia”, se trabaja a cubierto, los aparatos generan miles de resultados baratos que son fácilmente publicables. La taxonomía de campo es, en cambio, enormemente trabajosa, cara, necesita un largo aprendizaje y no se publica en revistas de alto impacto sino en tediosas monografías, revisiones, floras y faunas. ¿Quién eligiría la segunda vía?
Las puertas del ADN se abrieron y surgió una ola que dejó a los taxónomos sin fondos y sin espectativas. Como en tantas otras cosas, lo nuevo barrió lo clásico cuando, en realidad, el auténtico conocimiento, la comprensión de la realidad está aún en el campo. Rematan Wheeler y compañía:
“The need for this research has been masked, because molecular researchers could draw on centuries of banked morphology knowledge. That knowledge, however, is limited to a fraction of Earth’s species and will very soon be exhausted. Fashionable DNA bar-coding methods are a breakthrough for identification, but they will not supplant the need to formulate and rigorously test species hypotheses.”
Una noticia relacionada se reseña en el blog de Evolutionibus: una iniciativa que diseña “códigos de barras” genéticos para diferenciar las especies entre sí. Aparte de la idea, interesante, los tests para comprobar la bondad de la identificación de especies están basados, cómo no, en las especies descritas previa y convencionalmente. La presentación del proyecto, llamado The Barcode of Life Data System puede encontrarse en este PDF. Se trata, por tanto, de un proyecto de diagnóstico pero no de descubrimiento o de comprensión, entre otras cosas porque se basa sólo en la secuencia de unos cientos de nucleótidos de un único gen de ADN ribosómico. Por cierto que usan la taxonomía como comodín de una forma curiosa: si su test es conforme a las especies definidas con los métodos estándar es que funciona. Y si no funciona echan la culpa a la taxonomía convencional que no ha conseguido separar esas “especies ocultas”. Así cualquiera.
Para finalizar: ¿saldrá el especialista en ADN a las selvas de Bolivia a buscar nuevas especies o a entender mejor las relaciones a nivel de comunidad? No porque no es su papel pero tampoco deberíamos perder de vista que el taxónomo se convierte en una variedad en peligro de extinción y la pérdida de especies avanza a pasos de gigante.
P.S. Pueden ver un buen post sobre los nuevos filum en Diario de un copépodo

 han pasado por aquí
han pasado por aquí














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}